
Le tendon est un élément important de l’unité musculotendineuse. Il est l’intermédiaire entre les fibres musculaires et la surface osseuse.
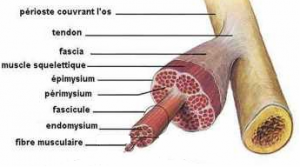
Les tendons sont de forme variable, ronds, plats, courts ou encore longs. Ils sont composés de différents faisceaux de fibres de collagène de type I et d’élastine entre lesquelles sont plaqués les ténocytes, cellules conjonctives spécialisées.
La masse totale d’un tendon comprend environ 30 % de collagène, 2 % d’élastine au sein d’une matrice extracellulaire (68 %) composée à 2/3 d’eau et 1/3 de cellules ténocytes et ténoblastes.
Les tendons ont une vascularisation pauvre et indépendante. L’apport sanguin autour et au sein du tendon lui-même est multiplié par 3 à 7 lors d’un exercice physique. Leur innervation sensitive est riche et intervient dans la régulation de la contraction musculaire. Le métabolisme est bas mais peut augmenter selon les sollicitations mécaniques.
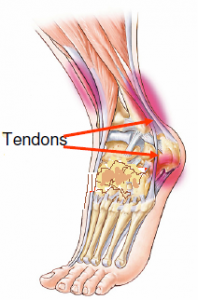
À quoi ressemble un tendon au microscope ?
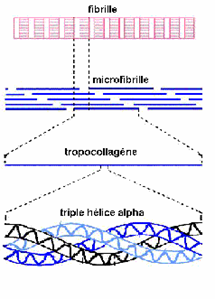
L’unité structurale du collagène est le tropcollagène, une protéine longue de 240 nm et large de 1,5 nm, qui forme la triple hélice de la molécule de collagène de type I en associant deux hélices a1 et une hélice a2 enrobées d’une fine couche de protéoglycanes et de glycosaminoglycanes. Une microfibrille de collagène est la réunion de cinq molécules de collagène. Une fibrille de collagène, d’environ 100 nm de diamètre est la réunion de plusieurs microfibrilles.
La fibre de collagène, de 1 à 20 μm de diamètre, union de plusieurs fibrilles de collagène est l’unité tendineuse la plus petite visible au microscope optique. Cette fibre de collagène comporte, entre les fibrilles, les ténocytes et quelques fibres d’élastine de 1 μm de longueur environ. Plusieurs fibres de collagène se rassemblent pour former les faisceaux fibreux primaires ou subfascicules.
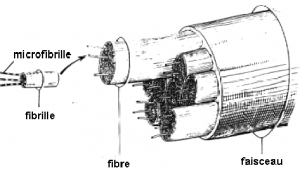
Les tendons de l’homme, quelles que soient leur forme et leurs dimensions, sont constitués de faisceaux fibreux primitifs ou primaires (subfascicule). Ceux-ci s’agencent pour former des faisceaux fibreux secondaires (fascicule) qui eux-mêmes se réunissent, donnant les faisceaux fibreux tertiaires. Les plus volumineux, comme le tendon calcanéen, sont constitués de plusieurs faisceaux tertiaires et sont alors nommés faisceaux quaternaires.
La substance nutritive extracellulaire enrobant le collagène et les ténocytes est composée de nombreuses molécules : protéoglycanes, glycosaminoglycanes, glycoprotéines… Une membrane fine, l’endotendon, contenant vaisseaux sanguins, lymphatiques et nerfs, entoure les faisceaux fibreux primaires, secondaires et tertiaires. L’unité tendineuse est recouverte par l’épitendon, gaine de tissu conjonctif lâche contenant les vaisseaux, lymphatiques et nerfs destinés au tendon.
Une troisième couche de tissu conjonctif, le paratendon, entoure encore le tendon. Cette membrane est parfois remplacée par une membrane synoviale. L’épitendon et le paratendon forment le péritendon, auquel on associe des propriétés tribologiques c’est à dire de glissement avec les tissus voisins. Ces structures conjonctives sont en continuité avec celles du corps musculaire constituant une véritable charpente conjonctive et fibreuse parcourant l’ensemble de l’unité musculotendineuse.
À quoi sert un tendon ?
Les muscles striés de la vie de relation présentent à leurs extrémités, des parties nacrées qui partagent avec eux des propriétés différentes d’élasticité. Les tendons sont constitués de faisceaux de nature fibreuse, denses, flexibles très résistants doués de propriétés viscoélastiques, capables d’emmagasiner de l’énergie pour la restituer secondairement et de s’adapter au cours de l’exercice physique.
Les tendons, permettent aux muscles de s’insérer sur les pièces squelettiques, ils peuvent être divisés en trois régions :
- La jonction musculotendineuse
- Le corps du tendon
- La jonction tendino-osseuse.
La jonction tendon-os est appelée enthèse. Elle peut être décomposée en quatre zones : la première correspond à une substance tendineuse classique, la deuxième à un fibrocartilage, la troisième à un fibrocartilage minéralisé et la quatrième à l’os proprement dit. Cette variation histologique permet une transition mécanique progressive en diffusant les forces de traction le long du tendon et en limitant les contraintes à l’insertion musculotendineuse.
La jonction myotendineuse transfert les forces musculaires au tendon et représente la zone de croissance musculaire. Les fibrilles de collagène du tendon sont insérées au fond de récessus formés par les myofibroblastes, ce qui permet de distribuer la tension générée par les protéines contractiles musculaires aux fibres tendineuses. Cette jonction est la zone la plus faible, mécaniquement, de l’unité musculotendineuse.
Les tendons jouent un rôle indispensable dans la régulation de la contraction musculaire, notamment grâce aux mécanorécepteurs de type III de Golgi, un organe tendineux spécifique. Sa situation à la jonction tendino-musculaire lui permet de recueillir des informations sur l’étirement musculaire et de coordonner ainsi l’activité musculaire.
Les tendons ne servent donc pas uniquement d’attaches anatomiques pour les muscles aux os, ils permettent aussi aux muscles de transférer des forces mécaniques aux articulations en amortissant leurs contraintes sur les structures périarticulaires : ligaments, capsules…

Quelle est la biologie du tendon ?
La consommation d’oxygène des tendons est comparable à celle des ligaments. Elle est estimée 7,5 fois moins importante que celle des muscles squelettiques. Le métabolisme tendineux est restreint mais sa capacité de produire de l’énergie en anaérobie est particulièrement développée. Ces caractéristiques rendent les tendons capables de supporter des poids et de résister à des tensions pendant de longues périodes en évitant l’ischémie et donc le risque de nécrose.
Néanmoins, ce taux métabolique bas rend la cicatrisation tendineuse lente. Au cours d’un exercice physique, la consommation d’oxygène est fonction des contraintes mécaniques exercées au sein du tendon. Elle est estimée 3 à 6 fois supérieure à la consommation de base.
Mécanique des tendons
Les tendons sont plus résistants que les muscles, ils sont soumis à des forces de tension et de compression qui peuvent supporter 17 fois le poids corporel total moyen d’un individu. Le tissu tendineux est capable de modifier sa structure, et sa composition est fonction des contraintes mécaniques qu’il subit.
L’entraînement physique induit des modifications mécaniques et biochimiques permettant de rendre plus résistant un tendon. Des travaux récents chez l’homme ont montré une augmentation du turnover du collagène de type I après exercice physique. Au cours d’un exercice physique de 60 minutes, la synthèse de collagène est augmentée de 100 % et se poursuit, de façon moindre, pendant 3 jours. Le turnover protéinique est également accéléré pendant 1 à 3 jours après l’exercice. L’effet de l’exercice physique peut devenir délétère lorsque les séances d’entraînement sont trop proches, le catabolisme devenant alors majoritaire. L’hypertrophie tendineuse et l’augmentation de la résistance mécanique du tendon résultent d’un entraînement régulier et progressif.
À l’inverse, un étirement excessif ou des contraintes déséquilibrées ont tendance à accroître la production de médiateurs de l’inflammation et à créer des lésions tendineuses aiguës ou chroniques.
Il existe donc une adaptation du tissu tendineux aux contraintes mécaniques ; cette mécanotransduction se fait tant à l’échelle macroscopique que moléculaire.
Les enthèses supportent des forces de tension, compression et de cisaillement pouvant être 4 fois supérieures à celles observées dans la portion moyenne du tendon.
La courbe typique de déformation d’un tendon par étirement montre une première phase, de croissance exponentielle qui correspond à la mise en tension progressive des fibres tendineuses.
À partir du moment où toutes les fibres tendineuses sont tendues, une seconde phase linéaire fait suite jusqu’à environ 4 % d’élongation (cette phase linéaire permet de calculer le module de Young du tendon, c’est-à-dire la pente de la courbe). La troisième portion de la courbe est extraphysiologique, elle va de 4 % d’élongation, limite maximale de rupture microscopique, à 10-14 % d’élongation, limite à laquelle on peut observer les premières lésions macroscopiques. Cette phase se poursuit par une dernière portion de courbe en plateau qui va jusqu’à la rupture tendineuse.




